Mẹo về What is the role of telomerase in end replication of chromosomes in eukaryotic cells? Chi Tiết
Lê Hoàng Hưng đang tìm kiếm từ khóa What is the role of telomerase in end replication of chromosomes in eukaryotic cells? được Update vào lúc : 2022-11-13 08:50:10 . Với phương châm chia sẻ Mẹo về trong nội dung bài viết một cách Chi Tiết Mới Nhất. Nếu sau khi Read tài liệu vẫn ko hiểu thì hoàn toàn có thể lại Comments ở cuối bài để Mình lý giải và hướng dẫn lại nha.Telomeres
Nội dung chính Show- IntroductionBudding Yeast, Saccharomyces cerevisiaeThe Need for TelomeresThe Epigenetic Regulation of Telomere Maintenance in Aging11 Telomeres as Epigenetic AgentsTelomere Maintenance in the Dynamic Nuclear ArchitectureAbbreviationsTelomeres, Aging, and NutritionMolecular Cell
BiologyTelomeric Repeat-Containing RNACrosstalk Between Non-Coding RNAs and the Epigenome in Development9.2.3.2 Telomere StabilityTelomeres in Health and DiseaseOxidative stress and antioxidants in elderly womenRole of telomere length in oxidative stress and antioxidant defense in elderly womenWhat is the role of telomerase in end replication of chromosomes in eukaryotic cells explain the process of end replication with the help of self explanatory diagram?What is the role of telomerase in replication?How does telomerase solve the end replication problem of eukaryotes?Why is telomerase necessary in eukaryotes?
D.M. Baird, in Encyclopedia of Gerontology (Second Edition), 2007
Introduction
Telomeres are essential chromosomal structures that cap the ends of linear eukaryotic chromosomes. Incomplete DNA replication every time cells divide results in a gradual loss of telomeric sequences in human somatic tissues. Telomere erosion ultimately results in the loss of telomeric function, which in turn triggers the cells to enter a non-dividing but biochemically active state termed replicative senescence. Telomere-dependent replicative senescence is considered to have evolved as a tumor suppressive mechanism; indeed, 85% of human malignancies express the enzyme telomerase that synthesizes telomere repeat sequences, preventing telomere loss and facilitating unlimited cell division. Senescent cells have the potential to interfere with the tissue microenvironment, and the accumulation of senescent cells is considered to contribute to age-related tissue deterioration and disease. Consistent with this hypothesis, telomere loss is observed as a function of age in human tissue, and this loss has been linked to specific disease phenotypes.
Read full chapter
URL: https://www.sciencedirect.com/science/article/pii/B0123708702001852
Telomeres
T.T. Chow, ... W.E. Wright, in Brenner's Encyclopedia of Genetics (Second Edition), 2013
Budding Yeast, Saccharomyces cerevisiae
Telomeres in budding yeast are bound by two distinct protein complexes: the CST complex that binds the G-overhang and the Rap1 complex that binds the DNA duplex. The CST complex forms as Cdc13 binds to G-overhangs and recruits two binding partners (Stn1 and Ten1). CST deficiency results in C-strand degradation and accumulation of G-overhangs. The CST complex also plays a role in telomere replication. In addition, Rap1, together with Rif1 and Rif2, binds the double-stranded telomeres and functions to regulate telomere length through a negative feedback loop. Increased binding of Rif1 and Rif2 telomeres inhibits telomerase recruitment and prevents further elongation. Disruption of Rap1 binding results in the lengthening of telomeric sequence and a faster turnover rate of the terminal repeats. Moreover, Rap1 can also recruit a second group of proteins – Sir2, Sir3, and Sir4, which enhances the formation of heterochromatin, telomere silencing, and influence positioning of the telomeres the nuclear periphery.
Read full chapter
URL: https://www.sciencedirect.com/science/article/pii/B9780123749840015230
Telomeres
J.R.S. Fincham, in Encyclopedia of Genetics, 2001
The Need for Telomeres
Telomeres are found the ends of chromosomes; they provide the answer to two problems of chromosome management. First, there has to be something to distinguish true chromosome ends from the accidental ends resulting from chromosome breakage. There is much evidence from studies of the effect of radiation on chromosomes that broken ends are prone to indiscriminate rejoining, with the possibility of segmental rearrangement. Presumably they provide substrates for double-stranded DNA ligase, and they may also be subject to erosion by exonuclease. True chromosome termini must be sealed in some way to protect them against these hazards.
Second, there has to be a way of completing DNA replication. DNA polymerase extends DNA strands from their 3′ ends, and so the two strands of double-stranded DNA are replicated in opposite directions. The synthesis of one follows the replication fork and can be continuous, but the synthesis of the other runs ‘backwards’ in piecemeal fashion, each piece having to be initiated afresh by an RNA primer that is subsequently removed. The consequence is that the DNA strand with a 5′ terminus cannot be fully replicated by the regular mechanism, since there is no 3′ end to prime the filling of the gap left by removal of the last RNA primer (Figure 1). Indeed, there is evidence that the 5′-terminated strand, already shorter, may be further shortened, leaving an even longer single-stranded 3′ ‘tail.’ So, without some additional end-replication mechanism, one would expect the chromosome to get a little shorter with each cycle of replication. Cell viability depends on the constant replenishment of terminal sequences.
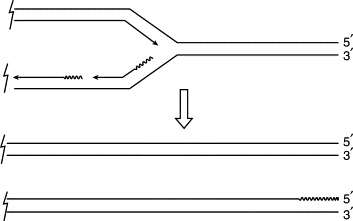
Figure 1. The end-replication problem. As the DNA replication fork progresses, one of the two parental strands can be replicated continuously, primed by its own 3′ end; the other (the lower in this figure) has to be replicated ‘backwards’ in patches, the replication of each patch being primed separately by primase RNA (shown here as a wavy line). When the replication fork reaches the chromosome end, the 5′-terminated strand of one daughter chromatid is incomplete, since the final RNA primer cannot be replaced by DNA.
Read full chapter
URL: https://www.sciencedirect.com/science/article/pii/B0122270800012763
The Epigenetic Regulation of Telomere Maintenance in Aging
Huda Adwan-Shekhidem, Gil Atzmon, in Epigenetics of Aging and Longevity, 2022
11 Telomeres as Epigenetic Agents
Telomeres are not only targets for epigenetic processes but also are chromosome domains acting themselves as epigenetic regulatory elements on other chromatin domains both in cis and trans positions. Telomere shortening affects the epigenetic status of telomeres and subtelomeres which in turn influence the telomere position effect (TPE). TPE is a phenomenon that refers to the ability of mammalian telomeres to silence subtelomeric genes. Telomere shortening in humans is associated with a decrease in TPE while TPE increases with telomere elongation [99]. Progressive loss of telomeres as a result of telomerase deficiency in mice showed a decrease in the density of heterochromatin marks in both telomeric and subtelomeric regions. Additionally, it showed an associated raise in acetylation of histone H3 and H4 suggesting that distal changes telomere ends may influence the epigenetic state of subtelomeric chromatin [100]. Furthermore, the loss of subtelomeric DNA methylation elucidated by telomere shortening results in telomere instability, and since telomere shortening attract telomerase, this may indicate that telomeres have specific marks recognizable by the telomerase complex.
Read full chapter
URL: https://www.sciencedirect.com/science/article/pii/B978012811060700005X
Telomere Maintenance in the Dynamic Nuclear Architecture
E. Micheli, ... S. Cacchione, in Chromatin Regulation and Dynamics, 2022
Abbreviations
ALT
Alternative lengthening of telomeres
APBALT-associated promyelocitic leukemia (PML) nuclear body toàn thân
ATMAtaxia telangiectasia mutated kinase
ATRATM and Rad3-related kinase
ATRXα-Thalassemia/mental retardation syndrome X-linked
ChIPChromatin immunoprecipitation
CSTCTC1-STN1-TEN1
CTCFCCCTC-binding factor
DAXXDeath domain–associated protein 6
DDRDNA damage response
D-loopDisplacement loop
DSBDouble-strand DNA break
GARGly/Arg-rich domain
γ-H2AXPhosphorylated H2AX on Ser-139
HDACHistone deacetylase
HP1Heterochromatin protein 1
HRHomologous recombination
ISG15Interferon-stimulated gene 15
MNaseMicrococcal nuclease
MRNMRE11/RAD50/NBS1 complex
NHEJNonhomologous end joining
PMLPromyelocytic leukemia
POT1Protection of telomeres 1
Rap1Repressor-activator protein 1
SIRSilent information regulator complex
SWI/SNFSWItch/sucrose nonfermentable complex
TADTopologically associated domain
TASTelomere-associated sequences
TER (or TR or TERC)Telomerase RNA component
TERRATelomeric repeat–containing RNA
TERTTelomerase reverse transcriptase
TIFTelomere dysfunction–induced focus
TIN2TRF1-interacting protein 2
T-loopTelomere loop
TPETelomere position effect
TPE-OLDTPE over long distances
TPP1TINT1/PIP1/PTOP1 complex
TRF1Telomeric repeat–binding factor 1
TRF2Telomeric repeat–binding factor 2
TRFHTRF homology domain
Read full chapter
URL: https://www.sciencedirect.com/science/article/pii/B9780128033951000137
Telomeres, Aging, and Nutrition
Varinderpal Dhillon, ... Michael Fenech, in Molecular Basis of Nutrition and Aging, 2022
Summary
Telomeres are nucleoprotein structures the end of each chromosome. The nucleic acid sequence of telomeres is a TTAGGG hexamer repeat. The number of hexamer repeats can vary greatly from very few to thousands of repeats with most reported telomere lengths varying between 4-11 kilobases in humans. Several proteins (TRF1, TRF2, POT1, TIN2, TPP1, RAP1 and Tankyrase) are associated with telomere DNA to form the shelterin complex which interacts with enzymes such as telomerase and other proteins required for proper telomere maintenance and function. Shortening of telomeres may increase excessively with age and lead to telomere-end-fusions, chromosomal instability and accelerated senescence. Telomeres may become dysfunctional if base damage and/or DNA strand breaks accumulate in the telomere sequence. Several lines of evidence indicate that telomere shortening, telomere base damage and telomere strand breaks may increase due to poor nutrition and that dietary intervention has the potential to improve telomere integrity.
Read full chapter
URL: https://www.sciencedirect.com/science/article/pii/B9780128018163000108
Molecular Cell Biology
J.J.-L. Chen, J.D. Podlevsky, in Encyclopedia of Cell Biology, 2022
Telomeric Repeat-Containing RNA
Telomeres were initially viewed as transcriptionally silent, packed into the higher-order heterochromatin structure. It was later discovered that the C-strand of the telomere is used as template for transcription of the long non-coding RNA, termed telomeric repeat-containing RNA (TERRA), which as the name suggests contains telomeric DNA repeat sequences. TERRA has been identified within distantly related species, ranging from vertebrates to yeasts to plants, and is actively transcribed from approximately 25% of the telomeres within a cell (Azzalin et al., 2007). Human TERRA is highly heterogeneous, varying in length from 100 nucleotides to more than 9000 nucleotides, while yeast TERRA has a more consistent length of approximately 400 nucleotides. The levels of TERRA transcribed are seemingly dependent on telomere length, with shorter telomeres promoting TERRA transcription and longer telomeres repressing expression (Iglesias et al., 2011; Arnoult et al., 2012). Shorter telomeres have fewer telomere-binding proteins that are responsible for transcriptional silencing of the telomere. TERRA appears to aid in telomerase recruitment and the increased levels of TERRA short telomeres increase the rate of telomere extension these eroded telomere ends (Cusanelli et al., 2013). Additional functions for TERRA have been proposed, which include preventing separate telomeres from interacting and supporting chromosome end stability.
Read full chapter
URL: https://www.sciencedirect.com/science/article/pii/B9780123944474100422
Crosstalk Between Non-Coding RNAs and the Epigenome in Development
M. Berdasco, M. Esteller, in Chromatin Regulation and Dynamics, 2022
9.2.3.2 Telomere Stability
Telomeres shorten after each mitotic cell division in adult somatic cells leading to cell senescence and apoptosis (see also Chapter 13). However, telomeres can be replenished by the telomerase reverse transcriptase enzyme, an enzyme that is very active in stem cells and also in tumor cells. The shelterin complex of proteins is known to regulate telomerase activity by binding to telomeres and inducing the formation of a t-loop, a cap structure of 300 bp single-stranded DNA that stabilizes the telomere, preventing the telomere ends from being recognized as potential break points by the DNA repair machinery [70]. Six proteins form the shelterin complex in humans: TRF1, TRF2, POT1, TIN2, TPP1, and RAP1 [71]. Telomere repeat-containing RNAs (TERRAs) are a group of lncRNAs transcribed the telomeres that actively participate in the regulation of telomere homeostasis and telomere function [58] through different mechanisms, including the stabilization of chromosome ends or the prevention of senescence induced by DNA damage response. TERRA transcripts are actively recruited to chromosome ends through interactions with stable components of the telomeric structure [59]. First, TERRA associates with the shelterin components TRF1 and TRF2 [59], but also with members of the epigenetic machinery including the heterochromatin protein 1 (HP1), the lysine 9-histone 3 methyltransferase SUV39H1 or the component of the NuA2 histone acetyltransferase complex, MORF4L2 [72,73]. Recent findings thus suggest that TERRA could act as a scaffold molecule promoting the binding of proteins associated with heterochromatin formation to the chromosome ends. Second, TERRA transcripts can base-pair with their template DNA strand, forming RNA:DNA hybrid structures known as R-loops telomere regions [60]. Interestingly, accumulation of telomeric R-loops promotes also the alternative lengthening of telomeres (ALT) pathway by facilitating homologous recombination between telomeric repeats of different chromosomes, and delays senescence in cells lacking telomerase [60]. Furthermore, tight regulation of the levels of TERRA during the cell cycle is involved in telomere protection and in the suppression of the DNA damage response telomeres [74]. During telomeric DNA replication, exposed ssDNA is thus bound by the ssDNA-binding protein RPA, which is essential for DNA replication and activation of the ATR checkpoint. RPA can be displaced by another ssDNA binding protein, hnRNAP1, which can be sequestered by TERRA [75]. TERRA expression changes during the cell cycle: it is expressed low levels in the late S phase while its expression increases in early G1 phase. As TERRA binds hnRNPA1, decreased TERRA during S phase facilitates hnRNPA1 binding to telomeric DNA and RPA displacement [75]. Release of RPA from telomeres prevents erroneous activation of ATR during cell cycle progression, enhancing telomere protection. Finally, it has been postulated that TERRA is also involved in the direct regulation of telomerase activity. In vitro assays showed that TERRA inhibits telomerase activity [76]; however, its regulatory role in vivo is still unclear.
Read full chapter
URL: https://www.sciencedirect.com/science/article/pii/B9780128033951000095
Telomeres in Health and Disease
Yoshiaki Sunami, ... Daniel Hartmann, in Progress in Molecular Biology and Translational Science, 2014
Abstract
Telomeres stabilize open chromosome ends and protect them against chromosomal end-to-end fusions, breakage, instability, and nonreciprocal translocations. Telomere dysfunction is known to lead to an impaired regenerative capacity of hepatocytes and an increased cirrhosis formation in the context of acute and chronic liver injury. In addition, telomere dysfunction and telomerase mutations have been associated with the induction of chromosomal instability and consequently with cirrhosis development and hepatocarcinogenesis. The identification of molecular mechanisms related to telomere dysfunction and telomerase activation might lead to new therapeutic strategies. In this chapter, we are reviewing the current knowledge about the importance of telomere dysfunction in liver diseases.
Read full chapter
URL: https://www.sciencedirect.com/science/article/pii/B9780123978981000074
Oxidative stress and antioxidants in elderly women
Brunna Cristina Bremer Boaventura, ... Francieli Cembranel, in Aging (Second Edition), 2022
Role of telomere length in oxidative stress and antioxidant defense in elderly women
Telomere maintenance might be a key factor in relation to the cumulative effects of genetic, environmental, and lifestyle factors involved in aging and aging-related diseases.16 Various nutrients influence telomere length via mechanisms that reflect their roles in cellular functions including DNA repair and chromosome maintenance, DNA methylation, inflammation, oxidative stress, and the activity of the enzyme telomerase (Fig. 3). Damage to telomeric DNA due to either oxidative stress or nucleotide precursors results in shorter telomeres.17 In this context, antioxidant nutrients can reduce the erosion of telomeres through the potential to influence the regulation of telomere length.17

Fig. 3. Potential mechanisms behind the influence of nutrients on telomere length. Dietary nutrients and oxidative stress influence telomere length by various mechanisms that reflect their role in cellular functions. Dashed line indicates effect of deficiency of a nutrient.
Reprinted from Paul L. Diet, nutrition and telomere length. J Nutr Biochem 2011;22(10):895–901, Copyright 2013, with permission from Elsevier.Although oxidative stress gene expression and telomere shortening may be epiphenomena of an underlying aging process, allelic variation in oxidative stress genotypes is fixed and may contribute to individual differences in both telomere length and biological aging.18 An investigation on elderly persons demonstrated that oxidative genes that are associated with shorter telomeres are also associated with the impairment of physical biomarkers of aging.18 The authors hypothesized that pathways involving altered Cu/Fe metabolism, glutathione transferase, and mitochondrial function are likely to be centered on sirtuins, providing a possible functional link between redox biology and telomere biology. Hence, associations between telomere length and physical biomarkers of aging may, in part, reflect cellular redox status as underlying common causes.18
The relationship between estimated endogenous estrogen exposure and telomere length was evaluated in a sample of postmenopausal women risk of cognitive decline.8 The authors found that greater endogenous estrogen exposure, as measured by a longer duration of reproductive years, was related to longer telomere length.
The telomerase molecule, which synthesizes telomeres, contains an estrogen-responsive element in its promoter region, which determines that telomerase activity is higher in females than in males.19 Longer telomeres in women have been ascribed to the ability of estrogen to upregulate telomerase and the same time reduce oxidative stress.8 In addition, it has been observed that telomerase is also regulated by glutathione.19
Read full chapter
URL: https://www.sciencedirect.com/science/article/pii/B9780128186985000146
What is the role of telomerase in end replication of chromosomes in eukaryotic cells explain the process of end replication with the help of self explanatory diagram?
Telomerase is a reverse transcriptase that adds telomeric repeats to the 3′ ends of each chromosome (3). In doing so, telomerase makes up for the shortcomings of semiconservative DNA replication, which cannot complete the synthesis of chromosome ends.What is the role of telomerase in replication?
Telomeres provide a mechanism for their replication by semiconservative DNA replication and length maintenance by telomerase. Through telomerase repression and induced telomere shortening, telomeres provide the means to regulate cellular life span.How does telomerase solve the end replication problem of eukaryotes?
Telomerase adds complementary RNA bases to the 3′ end of the DNA strand. Once the 3′ end of the lagging strand template is sufficiently elongated, DNA polymerase adds the complementary nucleotides to the ends of the chromosomes; thus, the ends of the chromosomes are replicated.Why is telomerase necessary in eukaryotes?
Over the years, it became clear that most eukaryotes use a telomerase based mechanism to replicate the very ends of chromosomes. This activity solves the first end replication problem because telomerase can elongate the 3' single strand G-tails in the absence of a DNA template. Tải thêm tài liệu liên quan đến nội dung bài viết What is the role of telomerase in end replication of chromosomes in eukaryotic cells?